
Volume 13, Issue 3 (10-2025)
Jorjani Biomed J 2025, 13(3): 25-30 |
Back to browse issues page


Download citation:
BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:



BibTeX | RIS | EndNote | Medlars | ProCite | Reference Manager | RefWorks
Send citation to:
Heydarzadeh E, Azizbeigi K, Mohamadzadeh Salamat K. Aerobic exercise and resveratrol cooperatively modulate hippocampal mitochondrial mitophagy in Alzheimer’s disease rat models. Jorjani Biomed J 2025; 13 (3) :25-30
URL: http://goums.ac.ir/jorjanijournal/article-1-1066-en.html
URL: http://goums.ac.ir/jorjanijournal/article-1-1066-en.html



1- Department of Physical Education, Sa.C., Islamic Azad University, Sanandaj, Iran
2- Department of Physical Education, Sa.C., Islamic Azad University, Sanandaj, Iran ,kazizbeigi@iau.ac.ir
2- Department of Physical Education, Sa.C., Islamic Azad University, Sanandaj, Iran ,
Full-Text [PDF 518 kb]
(1441 Downloads)
| Abstract (HTML) (5386 Views)
Resveratrol (20 mg/kg, Sigma-Aldrich) or an equivalent volume of saline was administered orally by gavage every morning (8:00-10:00 AM) for eight weeks (21).
Tissue collection and assay validation
Forty-eight hours after the last training session, and following 12-14 hours of fasting, the animals were anesthetized and sacrificed. Hippocampal tissue was isolated, washed in saline, placed into tubes containing RNA preservative, rapidly frozen in liquid nitrogen, and stored at –80°C until analysis. Table 1 presents the primer sequences used for PINK1, PARKIN, and GAPDH. Total RNA was extracted, reverse-transcribed into complementary DNA (cDNA), and analyzed by PCR to assess gene expression.
Real-time PCR
Twenty milligrams of tissue were minced and total cellular RNA was extracted using thiazole solution. cDNA synthesis was performed via reverse transcription. The cDNA was amplified using SYBR Green master mix (Thermo Scientific, USA) and the primers listed in Table 1. For mRNA quantification, 1 μg of total RNA was treated with RQ1 RNase-free DNase I enzyme (Promega) prior to reverse transcription. The thermal cycling protocol consisted of an initial denaturation at 95°C for 10 minutes, followed by 40 cycles of 20 seconds at 95°C, 30 seconds at 60°C, and 50 seconds at 72°C. Gene expression was evaluated using the comparative threshold cycle (Ct) method, normalized as (Control Ct) - (Target Ct) = ΔCt, and relative gene expression was calculated using 2-ΔCt.
Statistical analysis
Data distribution was assessed for normality using the Shapiro-Wilk test and for homogeneity of variances using Levene's test. Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test. Data were analyzed with SPSS version 26, and statistical significance was set at P ≤ 0.05.
Results
The average weight of the groups at baseline and after eight weeks is shown in Table 2.
At baseline, one‑way ANOVA revealed no significant differences in body weight among the groups (P-Value = 0.783). Paired t‑tests showed that after eight weeks, body weight increased significantly compared with baseline in the NO, AD, ADT, ADRSV, and ADTRSV groups (All P-Value = 0.001). Between‑group comparisons using one‑way ANOVA demonstrated that the ADTRSV group had a slightly but statistically significantly higher body weight than the NO group (P-Value = 0.043).
Data analysis using one-way ANOVA revealed significant differences in PINK1 expression among groups (F = 13.031, P-Value < 0.0001; Figure 2). Tukey's post-hoc tests demonstrated a significant decrease in PINK1 expression in the AD (P-Value < 0.0001), ADT (P-Value = 0.024), and ADRSV (P-Value = 0.032) groups compared to the NO group. A significant increase was observed in the ADT (P-Value = 0.043), ADRSV (P-Value = 0.033), and ADTRSV (P-Value < 0.0001) groups compared to the AD group. ADTRSV showed higher PINK1 levels than both ADT (P-Value = 0.036) and ADRSV (P-Value = 0.046) groups.
Similarly, one-way ANOVA revealed significant differences in PARKIN expression among groups (F = 16.851, P-Value < 0.0001; Figure 3). Tukey's post-hoc tests indicated a significant reduction in PARKIN expression in the AD (P-Value < 0.0001), ADT (P-Value = 0.006), and ADRSV (P-Value = 0.001) groups compared to the NO group. Significant increases were observed in the ADT (P-Value = 0.005), ADRSV (P-Value = 0.046), and ADTRSV (P-Value < 0.0001) groups compared to the AD group, with ADTRSV exhibiting higher PARKIN levels than the ADRSV group (P-Value = 0.046).
Discussion
This study demonstrated a significant reduction in PINK1 and PARKIN expression following Alzheimer’s disease (AD) induction, reinforcing the critical role of the PINK1-PARKIN pathway in mitochondrial homeostasis and AD pathology. While previous studies, such as Fang et al. (2019), reported impaired mitophagy in AD models (22), our findings provide direct evidence linking decreased PINK1 and PARKIN expression to hippocampal mitochondrial dysfunction, a region crucial for memory and cognition. This supports the hypothesis that mitophagy impairment contributes to the accumulation of damaged mitochondria, exacerbating neuronal dysfunction and death in AD.
Clinical studies by Castellazzi et al. (2019) also suggest that autophagy-related proteins such as ATG5 and PARKIN could serve as early diagnostic biomarkers for cognitive decline in elderly AD patients (23). Our findings align with these observations and extend them by demonstrating that both aerobic exercise and resveratrol (RSV) supplementation can restore PINK1 and PARKIN levels, potentially counteracting mitophagy deficits in AD. While some studies have reported limited effects of resveratrol on cognition in clinical populations - likely due to bioavailability, dosage, and duration - our results confirm its potential to restore mitophagy in AD. Additionally, some studies have reported limited cognitive effects of RSV in clinical populations, highlighting the need to consider factors such as bioavailability, dosage, and treatment duration as potential confounders (24,25).
AD pathology is characterized by the accumulation of beta-amyloid plaques and neurofibrillary tangles composed of hyperphosphorylated tau protein (26). Both Aβ and tau disrupt the PINK1-PARKIN pathway, increase ROS production, and damage mitochondrial membranes (27). Our findings corroborate this model by showing decreased PINK1 and PARKIN expression in AD rats, suggesting that impaired mitophagy is a key mechanism underlying mitochondrial abnormalities in AD. The PINK1-PARKIN pathway governs ubiquitin-dependent mitophagy, directing damaged mitochondria to lysosomal degradation and maintaining cellular homeostasis (28). Disruption of this pathway likely contributes to the accumulation of dysfunctional mitochondria, promoting oxidative stress and neuronal death. Alternative pathways, such as PGC-1α-mediated biogenesis, AMPK/mTOR signaling, and NRF2-regulated antioxidant pathways, may also play important roles (29).
Exercise has emerged as a robust modulator of mitochondrial quality control. Previous reports confirm that aerobic exercise enhances PINK1-PARKIN-dependent mitophagy, improving resistance to mitochondrial decline and cognitive impairments in AD models (30-33). Mechanistically, exercise-induced mitophagy enhancement involves activation of signaling pathways such as the SIRT1-FOXO1/3 axis, which mediates PINK1/PARKIN expression in the hippocampus (34,35). Another pathway implicated is the Irisin/FNDC5-PINK1/PARKIN axis, where resistance exercise upregulates Irisin and FNDC5, leading to enhanced mitophagy and reduced oxidative stress (36,37). Our study did not directly measure these mediators, representing a limitation. Future research should incorporate assessments of SIRT1, AMPK, and related pathways to elucidate the molecular mechanisms underlying exercise-induced mitophagy improvements.
Resveratrol supplementation also significantly increased PINK1 and PARKIN expression in AD rats. Resveratrol is documented to induce autophagy via SIRT1 activation and AMPK phosphorylation (38,39), and it reduces unfolded protein response and inflammatory mediators, improves learning, and enhances proteostasis by modulating the AMPK/SIRT1 pathway (40-43). The antioxidant transcription factor NRF2, targeted by resveratrol, also regulates autophagy and proteasome proteins, which decline with age. Collectively, these mechanisms suggest resveratrol acts through multiple converging pathways to promote mitochondrial quality control.
Importantly, our study revealed that the combination of aerobic exercise and resveratrol supplementation produced a more pronounced increase in PINK1 and PARKIN expression than either treatment alone. This synergistic effect aligns with reports that exercise and resveratrol together enhance SIRT1 expression and reduce pro-apoptotic factors such as P53 (44). Combined treatment also reduces Aβ oligomers and modulates neuronal autophagy more effectively than single interventions (45). These findings underscore the potential of multi-modal strategies targeting mitophagy to achieve greater therapeutic benefits in AD.
Despite these promising results, some conflicting evidence exists regarding the cognitive benefits of resveratrol in clinical trials, often attributed to bioavailability challenges, dosage variations, and differences in treatment duration (35,36). The precise molecular pathways by which exercise and resveratrol modulate mitophagy remain incompletely understood, with other signaling cascades such as AMPK/mTOR, PGC-1α, and inflammatory pathways likely involved (46). Additional limitations include the absence of direct measurements of SIRT1 and AMPK activity and the lack of behavioral or cognitive outcome assessment. The relatively small sample size further limits generalizability. Future studies should address these gaps by including behavioral assessments, larger cohorts, and longitudinal designs to evaluate long-term effects of these interventions on mitophagy and cognitive function. Exploring dose-response relationships and optimizing treatment regimens will be essential for clinical translation. Investigating the interplay between mitophagy and other cellular processes, such as inflammation and proteostasis, may further clarify the complex pathophysiology of AD and identify new therapeutic targets.
Conclusion
In conclusion, this study demonstrated that Alzheimer’s disease induction leads to a significant reduction in hippocampal PINK1 and PARKIN expression, indicating impaired mitophagy. Both aerobic exercise and resveratrol supplementation independently improved these indices, with their combination producing a synergistic effect. These results suggest that a multi-modal intervention combining exercise and resveratrol may effectively enhance mitophagy, maintain mitochondrial proteostasis, and contribute to the amelioration of AD pathology.
To build upon these findings, future research should:
- Directly measure key signaling mediators such as SIRT1 and AMPK to elucidate molecular mechanisms.
- Incorporate behavioral and cognitive assessments to link molecular changes with functional outcomes.
- Explore dose optimization, long-term effects, and translational potential in clinical or preclinical models.
- Investigate additional pathways influencing mitophagy and mitochondrial dynamics to develop comprehensive therapeutic strategies.
By addressing these areas, future studies can advance our understanding of mitophagy regulation in AD and facilitate the development of effective interventions to slow or prevent disease progression.
Acknowledgement
The authors gratefully acknowledge the Islamic Azad University, Sanandaj Branch, for administrative and laboratory support during this PhD dissertation project, which was conducted at personal expense.
Funding sources
This study was self-funded by the authors. No external funding or financial support was received from public, commercial, or not-for-profit agencies.
Ethical statement
All animal procedures were conducted in accordance with the guidelines of the National Institutes of Health (NIH) for the care and use of laboratory animals and were approved by the Research Ethics Committee of the Islamic Azad University, Ayatollah Amoli Branch (Approval code: IR.IAU.MAMOL.REC.1403.174).
Conflicts of interest
The authors declare that there are no conflicts of interest related to this study.
Author contributions
All authors contributed to the conception, design, execution, data analysis, and manuscript preparation. All authors read and approved the final version of the manuscript.
Data availability statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Full-Text: (760 Views)
Introduction
Alzheimer's disease (AD) is a progressive neurodegenerative disorder predominantly affecting the elderly, and is the most common cause of dementia worldwide (1). Clinically, AD is characterized by progressive memory loss, cognitive impairment, and functional decline. Pathologically, hallmark features include extracellular amyloid-beta (Aβ) plaques, intracellular neurofibrillary tangles composed of hyperphosphorylated tau, neuroinflammation, vascular amyloidosis, and widespread neuronal and synaptic degeneration (2). Mitochondria are dynamic organelles responsible for ATP generation and metabolic regulation, and are crucial for neuronal health and survival (3). In AD, mitochondrial dysfunction manifests as impaired oxidative phosphorylation, reduced ATP production, overproduction of reactive oxygen species (ROS), and disrupted calcium homeostasis, exacerbating synaptic failure and neuronal death (4,5). Mounting evidence highlights a “vicious cycle” in which mitochondrial disturbances promote Aβ deposition and tau phosphorylation (6). Additionally, mitochondrial dynamics (Fission and fusion) imbalance and reduced biogenesis further impair neuronal energy metabolism and resilience (7). Recent literature emphasizes that mitochondrial abnormalities, including oxidative stress and mitophagy dysfunction, are closely intertwined with AD pathology and accelerate neurodegeneration.
A key mechanism for maintaining mitochondrial integrity is mitochondrial proteostasis, which involves the mitochondrial unfolded protein response (UPRmt) and mitophagy, the selective removal of damaged mitochondria (2,3). UPRmt also activates SIRT3 and antioxidant enzymes as part of proteostasis responses. Mitophagy can occur basally, ensuring regular turnover under normal conditions, and can be induced in response to stress. Its regulation is governed by pathways including PINK1/PARKIN, AMPK, and SIRT1 (5). Basal mitophagy supports homeostasis, while induced mitophagy protects neurons from mitochondrial insults. However, the interplay among mitophagy, mitochondrial biogenesis, and dynamics remains poorly understood in AD models, constituting a major gap in knowledge (8).
Non-pharmacological interventions such as aerobic exercise and resveratrol (RSV) have gained attention for modulating mitochondrial function (9,10). Exercise activates AMPK/NAD+-dependent signaling, promotes mitochondrial biogenesis via PGC-1α, and improves dynamics (11,12). Studies in Parkinson’s disease models demonstrate that treadmill running improves mitochondrial dysfunction by increasing mitophagy markers such as PINK1, PARKIN, and p62 (13). RSV, a polyphenol, exhibits antioxidant and anti-inflammatory properties, enhances biogenesis via AMPK/PGC-1α, and activates mitophagy through the PINK1–PARKIN and SIRT1 pathways (14). However, clinical translation is limited by RSV’s poor bioavailability, rapid metabolism, and limited blood-brain barrier penetration; novel delivery approaches are being explored (15). Clinical findings remain inconsistent due to variability in dosage, duration, study design, and population heterogeneity (9). Some trials report cognitive benefits and reduced Aβ pathology, while others show minimal effects, highlighting ongoing controversies. An animal study combining aerobic training and RSV demonstrated synergistic activation of the AMPK/PGC-1α/SIRT1 pathway in AD models, with combined treatment outperforming individual interventions (16). Despite this evidence, no prior studies have comprehensively examined the combined effects of exercise and RSV on multiple mitochondrial quality-control mechanisms - mitophagy, biogenesis, and dynamics - in AD models. Addressing this gap holds significant clinical relevance, as targeting several aspects of mitochondrial health may better preserve cognition and delay AD progression (17).
Therefore, this study aimed to explore the synergistic effects of aerobic exercise and RSV on mitophagy in the hippocampal tissue of AD rats. We hypothesized that combined treatment would synergistically enhance mitophagy, mitochondrial biogenesis, and dynamics via the SIRT1/PINK1/PARKIN signaling pathway, ultimately improving mitochondrial function and cognitive performance.
Methods
This laboratory-based experimental study was conducted in accordance with animal protection policies outlined by the Helsinki Convention and the National Institutes of Health (NIH) guidelines for the care and use of laboratory animals. The experimental protocol was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of Islamic Azad University, Ayatollah Amoli Branch (Approval code: IR.IAU.MAMOL.REC.1403.174).
Thirty-five 8-week-old male Wistar rats (Average weight 223.17 ± 9.08 g) were selected, as this age corresponds to young adulthood in rats and is suitable for modeling early-stage Alzheimer’s disease. Male rats were chosen to avoid hormonal fluctuations associated with the estrous cycle that could affect experimental outcomes. All rats underwent veterinary screening to confirm good health and the absence of pre-existing conditions. Rats were housed under controlled conditions (22 ± 3°C, 12:12-hour light-dark cycle) with ad libitum access to water and a standardized rodent diet (20% protein, 5% fat, 55% carbohydrate, and essential vitamins and minerals). The sample size of seven rats per group was determined based on prior studies and calculated to achieve a statistical power of 80% with an alpha level of 0.05 to detect significant differences in mitochondrial autophagy markers.
After a one-week acclimatization period-including daily handling and habituation to the treadmill apparatus to reduce stress and improve exercise compliance-the rats were randomly assigned into five groups of seven rats each: (1) Normal (NO), (2) Alzheimer’s (AD), (3) Alzheimer’s-Training (ADT), (4) Alzheimer’s-Resveratrol (ADRSV), and (5) Alzheimer’s-Training-Resveratrol (ADTRSV). Randomization was performed using a computer-generated random number sequence.
Alzheimer’s induction
Amyloid-β 1-42 (Sigma-Aldrich) was dissolved in double-distilled sterilized water and incubated at 37°C for one week to promote aggregation. The solution was stored at 4°C and used within 24 hours to maintain peptide integrity. Rats were anesthetized with ketamine (50 mg/kg) and xylazine (5 mg/kg), and placed in a stereotaxic apparatus. The scalp was shaved, and bregma and lambda sutures were identified via sagittal incision. Stereotaxic coordinates for the CA1 hippocampal region were AP –3.6 mm, ML ±2.0 mm, DV –2.8 mm from bregma. Using a Hamilton syringe, 2 μL of amyloid-β solution was injected into the hippocampus over one minute. The needle was left in place for five minutes to prevent backflow before withdrawal (18).
Exercise protocol
The exercise regimen, as shown in Figure 1, commenced at two months of age and included two stages: familiarization (2 weeks) and adaptation to the full training protocol (8 weeks). During familiarization, rats performed treadmill exercises at 6-18 meters per minute for 15-45 minutes, five sessions in the first week, corresponding to moderate-intensity exercise. This speed corresponds to moderate-intensity exercise for Wistar rats, approximately 60-70% of their maximal oxygen uptake (VO2max), and is effective in inducing mitochondrial adaptations without causing excessive fatigue (19). The subsequent 8-week period involved running at 18 meters per minute for 45 minutes, five days per week (20).
Alzheimer's disease (AD) is a progressive neurodegenerative disorder predominantly affecting the elderly, and is the most common cause of dementia worldwide (1). Clinically, AD is characterized by progressive memory loss, cognitive impairment, and functional decline. Pathologically, hallmark features include extracellular amyloid-beta (Aβ) plaques, intracellular neurofibrillary tangles composed of hyperphosphorylated tau, neuroinflammation, vascular amyloidosis, and widespread neuronal and synaptic degeneration (2). Mitochondria are dynamic organelles responsible for ATP generation and metabolic regulation, and are crucial for neuronal health and survival (3). In AD, mitochondrial dysfunction manifests as impaired oxidative phosphorylation, reduced ATP production, overproduction of reactive oxygen species (ROS), and disrupted calcium homeostasis, exacerbating synaptic failure and neuronal death (4,5). Mounting evidence highlights a “vicious cycle” in which mitochondrial disturbances promote Aβ deposition and tau phosphorylation (6). Additionally, mitochondrial dynamics (Fission and fusion) imbalance and reduced biogenesis further impair neuronal energy metabolism and resilience (7). Recent literature emphasizes that mitochondrial abnormalities, including oxidative stress and mitophagy dysfunction, are closely intertwined with AD pathology and accelerate neurodegeneration.
A key mechanism for maintaining mitochondrial integrity is mitochondrial proteostasis, which involves the mitochondrial unfolded protein response (UPRmt) and mitophagy, the selective removal of damaged mitochondria (2,3). UPRmt also activates SIRT3 and antioxidant enzymes as part of proteostasis responses. Mitophagy can occur basally, ensuring regular turnover under normal conditions, and can be induced in response to stress. Its regulation is governed by pathways including PINK1/PARKIN, AMPK, and SIRT1 (5). Basal mitophagy supports homeostasis, while induced mitophagy protects neurons from mitochondrial insults. However, the interplay among mitophagy, mitochondrial biogenesis, and dynamics remains poorly understood in AD models, constituting a major gap in knowledge (8).
Non-pharmacological interventions such as aerobic exercise and resveratrol (RSV) have gained attention for modulating mitochondrial function (9,10). Exercise activates AMPK/NAD+-dependent signaling, promotes mitochondrial biogenesis via PGC-1α, and improves dynamics (11,12). Studies in Parkinson’s disease models demonstrate that treadmill running improves mitochondrial dysfunction by increasing mitophagy markers such as PINK1, PARKIN, and p62 (13). RSV, a polyphenol, exhibits antioxidant and anti-inflammatory properties, enhances biogenesis via AMPK/PGC-1α, and activates mitophagy through the PINK1–PARKIN and SIRT1 pathways (14). However, clinical translation is limited by RSV’s poor bioavailability, rapid metabolism, and limited blood-brain barrier penetration; novel delivery approaches are being explored (15). Clinical findings remain inconsistent due to variability in dosage, duration, study design, and population heterogeneity (9). Some trials report cognitive benefits and reduced Aβ pathology, while others show minimal effects, highlighting ongoing controversies. An animal study combining aerobic training and RSV demonstrated synergistic activation of the AMPK/PGC-1α/SIRT1 pathway in AD models, with combined treatment outperforming individual interventions (16). Despite this evidence, no prior studies have comprehensively examined the combined effects of exercise and RSV on multiple mitochondrial quality-control mechanisms - mitophagy, biogenesis, and dynamics - in AD models. Addressing this gap holds significant clinical relevance, as targeting several aspects of mitochondrial health may better preserve cognition and delay AD progression (17).
Therefore, this study aimed to explore the synergistic effects of aerobic exercise and RSV on mitophagy in the hippocampal tissue of AD rats. We hypothesized that combined treatment would synergistically enhance mitophagy, mitochondrial biogenesis, and dynamics via the SIRT1/PINK1/PARKIN signaling pathway, ultimately improving mitochondrial function and cognitive performance.
Methods
This laboratory-based experimental study was conducted in accordance with animal protection policies outlined by the Helsinki Convention and the National Institutes of Health (NIH) guidelines for the care and use of laboratory animals. The experimental protocol was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of Islamic Azad University, Ayatollah Amoli Branch (Approval code: IR.IAU.MAMOL.REC.1403.174).
Thirty-five 8-week-old male Wistar rats (Average weight 223.17 ± 9.08 g) were selected, as this age corresponds to young adulthood in rats and is suitable for modeling early-stage Alzheimer’s disease. Male rats were chosen to avoid hormonal fluctuations associated with the estrous cycle that could affect experimental outcomes. All rats underwent veterinary screening to confirm good health and the absence of pre-existing conditions. Rats were housed under controlled conditions (22 ± 3°C, 12:12-hour light-dark cycle) with ad libitum access to water and a standardized rodent diet (20% protein, 5% fat, 55% carbohydrate, and essential vitamins and minerals). The sample size of seven rats per group was determined based on prior studies and calculated to achieve a statistical power of 80% with an alpha level of 0.05 to detect significant differences in mitochondrial autophagy markers.
After a one-week acclimatization period-including daily handling and habituation to the treadmill apparatus to reduce stress and improve exercise compliance-the rats were randomly assigned into five groups of seven rats each: (1) Normal (NO), (2) Alzheimer’s (AD), (3) Alzheimer’s-Training (ADT), (4) Alzheimer’s-Resveratrol (ADRSV), and (5) Alzheimer’s-Training-Resveratrol (ADTRSV). Randomization was performed using a computer-generated random number sequence.
Alzheimer’s induction
Amyloid-β 1-42 (Sigma-Aldrich) was dissolved in double-distilled sterilized water and incubated at 37°C for one week to promote aggregation. The solution was stored at 4°C and used within 24 hours to maintain peptide integrity. Rats were anesthetized with ketamine (50 mg/kg) and xylazine (5 mg/kg), and placed in a stereotaxic apparatus. The scalp was shaved, and bregma and lambda sutures were identified via sagittal incision. Stereotaxic coordinates for the CA1 hippocampal region were AP –3.6 mm, ML ±2.0 mm, DV –2.8 mm from bregma. Using a Hamilton syringe, 2 μL of amyloid-β solution was injected into the hippocampus over one minute. The needle was left in place for five minutes to prevent backflow before withdrawal (18).
Exercise protocol
The exercise regimen, as shown in Figure 1, commenced at two months of age and included two stages: familiarization (2 weeks) and adaptation to the full training protocol (8 weeks). During familiarization, rats performed treadmill exercises at 6-18 meters per minute for 15-45 minutes, five sessions in the first week, corresponding to moderate-intensity exercise. This speed corresponds to moderate-intensity exercise for Wistar rats, approximately 60-70% of their maximal oxygen uptake (VO2max), and is effective in inducing mitochondrial adaptations without causing excessive fatigue (19). The subsequent 8-week period involved running at 18 meters per minute for 45 minutes, five days per week (20).
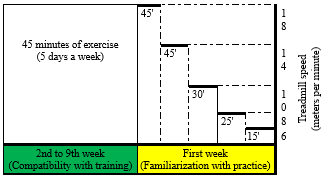 Figure 1. Exercise protocol |
Resveratrol (20 mg/kg, Sigma-Aldrich) or an equivalent volume of saline was administered orally by gavage every morning (8:00-10:00 AM) for eight weeks (21).
Tissue collection and assay validation
Forty-eight hours after the last training session, and following 12-14 hours of fasting, the animals were anesthetized and sacrificed. Hippocampal tissue was isolated, washed in saline, placed into tubes containing RNA preservative, rapidly frozen in liquid nitrogen, and stored at –80°C until analysis. Table 1 presents the primer sequences used for PINK1, PARKIN, and GAPDH. Total RNA was extracted, reverse-transcribed into complementary DNA (cDNA), and analyzed by PCR to assess gene expression.
Real-time PCR
Twenty milligrams of tissue were minced and total cellular RNA was extracted using thiazole solution. cDNA synthesis was performed via reverse transcription. The cDNA was amplified using SYBR Green master mix (Thermo Scientific, USA) and the primers listed in Table 1. For mRNA quantification, 1 μg of total RNA was treated with RQ1 RNase-free DNase I enzyme (Promega) prior to reverse transcription. The thermal cycling protocol consisted of an initial denaturation at 95°C for 10 minutes, followed by 40 cycles of 20 seconds at 95°C, 30 seconds at 60°C, and 50 seconds at 72°C. Gene expression was evaluated using the comparative threshold cycle (Ct) method, normalized as (Control Ct) - (Target Ct) = ΔCt, and relative gene expression was calculated using 2-ΔCt.
Statistical analysis
Data distribution was assessed for normality using the Shapiro-Wilk test and for homogeneity of variances using Levene's test. Statistical analysis was performed using one-way analysis of variance (ANOVA) followed by Tukey’s post-hoc test. Data were analyzed with SPSS version 26, and statistical significance was set at P ≤ 0.05.
|
Table 1. Primer pattern of PINK1 and PARKIN
 |
Results
The average weight of the groups at baseline and after eight weeks is shown in Table 2.
At baseline, one‑way ANOVA revealed no significant differences in body weight among the groups (P-Value = 0.783). Paired t‑tests showed that after eight weeks, body weight increased significantly compared with baseline in the NO, AD, ADT, ADRSV, and ADTRSV groups (All P-Value = 0.001). Between‑group comparisons using one‑way ANOVA demonstrated that the ADTRSV group had a slightly but statistically significantly higher body weight than the NO group (P-Value = 0.043).
Data analysis using one-way ANOVA revealed significant differences in PINK1 expression among groups (F = 13.031, P-Value < 0.0001; Figure 2). Tukey's post-hoc tests demonstrated a significant decrease in PINK1 expression in the AD (P-Value < 0.0001), ADT (P-Value = 0.024), and ADRSV (P-Value = 0.032) groups compared to the NO group. A significant increase was observed in the ADT (P-Value = 0.043), ADRSV (P-Value = 0.033), and ADTRSV (P-Value < 0.0001) groups compared to the AD group. ADTRSV showed higher PINK1 levels than both ADT (P-Value = 0.036) and ADRSV (P-Value = 0.046) groups.
Similarly, one-way ANOVA revealed significant differences in PARKIN expression among groups (F = 16.851, P-Value < 0.0001; Figure 3). Tukey's post-hoc tests indicated a significant reduction in PARKIN expression in the AD (P-Value < 0.0001), ADT (P-Value = 0.006), and ADRSV (P-Value = 0.001) groups compared to the NO group. Significant increases were observed in the ADT (P-Value = 0.005), ADRSV (P-Value = 0.046), and ADTRSV (P-Value < 0.0001) groups compared to the AD group, with ADTRSV exhibiting higher PARKIN levels than the ADRSV group (P-Value = 0.046).
|
Table 2. Comparison of average body weight among groups at baseline and after 8 weeks
 NO: Normal, AD: Alzheimer, ADT: Alzheimer-Training, ADRSV: Alzheimer-Resveratrol, ADTRSV: Alzheimer-Training-Resveratrol. * Difference from First week same group, c Difference from ADRSV group. 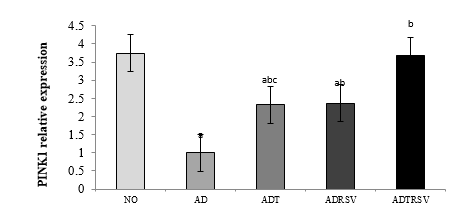 Figure 2. Expression levels of hippocampal PINK1 in different groups. a- Indicates significant decrease with the NO group; b- Indicates a significant increase with the AD group; c- Indicates a significant decrease with the ADTRSV group. Abbreviations: NO: Normal group; AD: Alzheimer group; ADT: Alzheimer-Training group; ADRSV: Alzheimer-Resveratrol group; ADTRSV: Alzheimer-Training-Resveratrol group. 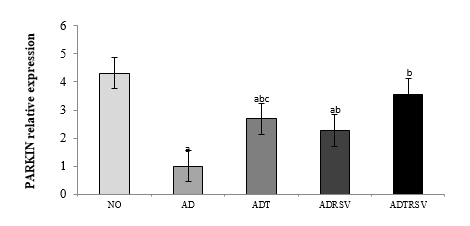 Figure 3. Expression levels of hippocampal PARKIN in different groups. a- Indicates significant decrease with the NO group; b- Indicates a significant increase with the AD group; c- Indicates a significant decrease with the ADTRSV group. Abbreviations: NO: Normal group; AD: Alzheimer group; ADT: Alzheimer-Training group; ADRSV: Alzheimer-Resveratrol group; ADTRSV: Alzheimer-Training-Resveratrol group. |
Discussion
This study demonstrated a significant reduction in PINK1 and PARKIN expression following Alzheimer’s disease (AD) induction, reinforcing the critical role of the PINK1-PARKIN pathway in mitochondrial homeostasis and AD pathology. While previous studies, such as Fang et al. (2019), reported impaired mitophagy in AD models (22), our findings provide direct evidence linking decreased PINK1 and PARKIN expression to hippocampal mitochondrial dysfunction, a region crucial for memory and cognition. This supports the hypothesis that mitophagy impairment contributes to the accumulation of damaged mitochondria, exacerbating neuronal dysfunction and death in AD.
Clinical studies by Castellazzi et al. (2019) also suggest that autophagy-related proteins such as ATG5 and PARKIN could serve as early diagnostic biomarkers for cognitive decline in elderly AD patients (23). Our findings align with these observations and extend them by demonstrating that both aerobic exercise and resveratrol (RSV) supplementation can restore PINK1 and PARKIN levels, potentially counteracting mitophagy deficits in AD. While some studies have reported limited effects of resveratrol on cognition in clinical populations - likely due to bioavailability, dosage, and duration - our results confirm its potential to restore mitophagy in AD. Additionally, some studies have reported limited cognitive effects of RSV in clinical populations, highlighting the need to consider factors such as bioavailability, dosage, and treatment duration as potential confounders (24,25).
AD pathology is characterized by the accumulation of beta-amyloid plaques and neurofibrillary tangles composed of hyperphosphorylated tau protein (26). Both Aβ and tau disrupt the PINK1-PARKIN pathway, increase ROS production, and damage mitochondrial membranes (27). Our findings corroborate this model by showing decreased PINK1 and PARKIN expression in AD rats, suggesting that impaired mitophagy is a key mechanism underlying mitochondrial abnormalities in AD. The PINK1-PARKIN pathway governs ubiquitin-dependent mitophagy, directing damaged mitochondria to lysosomal degradation and maintaining cellular homeostasis (28). Disruption of this pathway likely contributes to the accumulation of dysfunctional mitochondria, promoting oxidative stress and neuronal death. Alternative pathways, such as PGC-1α-mediated biogenesis, AMPK/mTOR signaling, and NRF2-regulated antioxidant pathways, may also play important roles (29).
Exercise has emerged as a robust modulator of mitochondrial quality control. Previous reports confirm that aerobic exercise enhances PINK1-PARKIN-dependent mitophagy, improving resistance to mitochondrial decline and cognitive impairments in AD models (30-33). Mechanistically, exercise-induced mitophagy enhancement involves activation of signaling pathways such as the SIRT1-FOXO1/3 axis, which mediates PINK1/PARKIN expression in the hippocampus (34,35). Another pathway implicated is the Irisin/FNDC5-PINK1/PARKIN axis, where resistance exercise upregulates Irisin and FNDC5, leading to enhanced mitophagy and reduced oxidative stress (36,37). Our study did not directly measure these mediators, representing a limitation. Future research should incorporate assessments of SIRT1, AMPK, and related pathways to elucidate the molecular mechanisms underlying exercise-induced mitophagy improvements.
Resveratrol supplementation also significantly increased PINK1 and PARKIN expression in AD rats. Resveratrol is documented to induce autophagy via SIRT1 activation and AMPK phosphorylation (38,39), and it reduces unfolded protein response and inflammatory mediators, improves learning, and enhances proteostasis by modulating the AMPK/SIRT1 pathway (40-43). The antioxidant transcription factor NRF2, targeted by resveratrol, also regulates autophagy and proteasome proteins, which decline with age. Collectively, these mechanisms suggest resveratrol acts through multiple converging pathways to promote mitochondrial quality control.
Importantly, our study revealed that the combination of aerobic exercise and resveratrol supplementation produced a more pronounced increase in PINK1 and PARKIN expression than either treatment alone. This synergistic effect aligns with reports that exercise and resveratrol together enhance SIRT1 expression and reduce pro-apoptotic factors such as P53 (44). Combined treatment also reduces Aβ oligomers and modulates neuronal autophagy more effectively than single interventions (45). These findings underscore the potential of multi-modal strategies targeting mitophagy to achieve greater therapeutic benefits in AD.
Despite these promising results, some conflicting evidence exists regarding the cognitive benefits of resveratrol in clinical trials, often attributed to bioavailability challenges, dosage variations, and differences in treatment duration (35,36). The precise molecular pathways by which exercise and resveratrol modulate mitophagy remain incompletely understood, with other signaling cascades such as AMPK/mTOR, PGC-1α, and inflammatory pathways likely involved (46). Additional limitations include the absence of direct measurements of SIRT1 and AMPK activity and the lack of behavioral or cognitive outcome assessment. The relatively small sample size further limits generalizability. Future studies should address these gaps by including behavioral assessments, larger cohorts, and longitudinal designs to evaluate long-term effects of these interventions on mitophagy and cognitive function. Exploring dose-response relationships and optimizing treatment regimens will be essential for clinical translation. Investigating the interplay between mitophagy and other cellular processes, such as inflammation and proteostasis, may further clarify the complex pathophysiology of AD and identify new therapeutic targets.
Conclusion
In conclusion, this study demonstrated that Alzheimer’s disease induction leads to a significant reduction in hippocampal PINK1 and PARKIN expression, indicating impaired mitophagy. Both aerobic exercise and resveratrol supplementation independently improved these indices, with their combination producing a synergistic effect. These results suggest that a multi-modal intervention combining exercise and resveratrol may effectively enhance mitophagy, maintain mitochondrial proteostasis, and contribute to the amelioration of AD pathology.
To build upon these findings, future research should:
- Directly measure key signaling mediators such as SIRT1 and AMPK to elucidate molecular mechanisms.
- Incorporate behavioral and cognitive assessments to link molecular changes with functional outcomes.
- Explore dose optimization, long-term effects, and translational potential in clinical or preclinical models.
- Investigate additional pathways influencing mitophagy and mitochondrial dynamics to develop comprehensive therapeutic strategies.
By addressing these areas, future studies can advance our understanding of mitophagy regulation in AD and facilitate the development of effective interventions to slow or prevent disease progression.
Acknowledgement
The authors gratefully acknowledge the Islamic Azad University, Sanandaj Branch, for administrative and laboratory support during this PhD dissertation project, which was conducted at personal expense.
Funding sources
This study was self-funded by the authors. No external funding or financial support was received from public, commercial, or not-for-profit agencies.
Ethical statement
All animal procedures were conducted in accordance with the guidelines of the National Institutes of Health (NIH) for the care and use of laboratory animals and were approved by the Research Ethics Committee of the Islamic Azad University, Ayatollah Amoli Branch (Approval code: IR.IAU.MAMOL.REC.1403.174).
Conflicts of interest
The authors declare that there are no conflicts of interest related to this study.
Author contributions
All authors contributed to the conception, design, execution, data analysis, and manuscript preparation. All authors read and approved the final version of the manuscript.
Data availability statement
The datasets generated and/or analyzed during the current study are available from the corresponding author upon reasonable request.
Editorial: Original article |
Subject:
Health
Received: 2025/03/9 | Accepted: 2025/09/10 | Published: 2025/09/23
Received: 2025/03/9 | Accepted: 2025/09/10 | Published: 2025/09/23
References
1. Breijyeh Z, Karaman R. Comprehensive review on Alzheimer’s disease: Causes and treatment. Molecules. 2020;25(24):5789. [View at Publisher] [DOI] [PMID] [Google Scholar]
2. Xu Y, Zheng F, Zhong Q, Zhu Y. Ketogenic diet as a promising non-drug intervention for Alzheimer's disease: Mechanisms and clinical implications. J Alzheimers Dis . 2023;92(4):1173-98. [View at Publisher] [DOI] [PMID] [Google Scholar]
3. Papa S, Martino PL, Capitanio G, Gaballo A, De Rasmo D, Signorile A, et al. The oxidative phosphorylation system in mammalian mitochondria. Adv Exp Med Biol. 2012:942:3-37. [View at Publisher] [DOI] [PMID] [Google Scholar]
4. Garaeva AA, Kovaleva IE, Chumakov PM, Evstafieva AG. Mitochondrial dysfunction induces SESN 2 gene expression through activating transcription factor 4. Cell Cycle . 2016;15(1):64-71. [View at Publisher] [DOI] [PMID] [Google Scholar]
5. Pickles S, Vigié P, Youle RJ. Mitophagy and quality control mechanisms in mitochondrial maintenance. Curr Biol. 2018;28(4):R170-R85. [View at Publisher] [DOI] [PMID] [Google Scholar]
6. Li X, Wu Z, Si X, Li J, Wu G, Wang M. The role of mitochondrial dysfunction in the pathogenesis of Alzheimer's disease and future strategies for targeted therapy. Eur J Med Res . 2025;30(1):434. [View at Publisher] [DOI] [PMID] [Google Scholar]
7. Wang S, Liao Z, Zhang Q, Han X, Liu C, Wang J. Mitochondrial dysfunction in Alzheimer's disease: a key frontier for future targeted therapies. Front Immunol . 2025:15:1484373. [View at Publisher] [DOI] [PMID] [Google Scholar]
8. Bar-Ziv R, Dutta N, Hruby A, Sukarto E, Averbukh M, Alcala A, et al. Glial-derived mitochondrial signals affect neuronal proteostasis and aging. Sci Adv. 2023;9(41):eadi1411. [View at Publisher] [DOI] [PMID] [Google Scholar]
9. Turner RS, Thomas RG, Craft S, Van Dyck CH, Mintzer J, Reynolds BA, et al. A randomized, double-blind, placebo-controlled trial of resveratrol for Alzheimer disease. Neurology. 2015;85(16):1383-91. [View at Publisher] [DOI] [PMID] [Google Scholar]
10. Van Acker ZP, Leroy T, Annaert W. Mitochondrial dysfunction, cause or consequence in neurodegenerative diseases? Bioessays. 2025;47(1):e2400023. [View at Publisher] [DOI] [PMID] [Google Scholar]
11. Kang J, Liu M, Yang Q, Dang X, Li Q, Wang T, et al. Exercise training exerts beneficial effects on Alzheimer's disease through multiple signaling pathways. Front Aging Neurosci . 2025:17:1558078. [View at Publisher] [DOI] [PMID] [Google Scholar]
12. De la Rosa A, Olaso-Gonzalez G, Arc-Chagnaud C, Millan F, Salvador-Pascual A, García-Lucerga C, et al. Physical exercise in the prevention and treatment of Alzheimer's disease. J Sport Health Sci. 2020;9(5):394-404. [View at Publisher] [DOI] [PMID] [Google Scholar]
13. O'Reilly CL, Miller BF, Lewis Jr TL. Exercise and mitochondrial remodeling to prevent age-related neurodegeneration. J Appl Physiol (1985). 2023;134(1):181-9. [View at Publisher] [DOI] [PMID] [Google Scholar]
14. Chandra G, Kundu M, Rangasamy SB, Dasarathy S, Ghosh S, Watson R, et al. Increase in mitochondrial biogenesis in neuronal cells by RNS60, a physically-modified saline, via phosphatidylinositol 3-kinase-mediated upregulation of PGC1α. J Neuroimmune Pharmacol. 2018;13(2):143-62. [View at Publisher] [DOI] [PMID] [Google Scholar]
15. Puranik N, Kumari M, Tiwari S, Dhakal T, Song M. Resveratrol as a Therapeutic Agent in Alzheimer's Disease: Evidence from Clinical Studies. Nutrients. 2025;17(15):2557. [View at Publisher] [DOI] [PMID] [Google Scholar]
16. Rashet A, Abdi A, Barari A. Synergistic Role of Aerobic Training and Resveratrol on AMPK/PGC1-α/SIRT1 Pathway in the Hippocampus of Rats with Alzheimer's Disease. J Arch Mil Med. 2024;12(1):e144281. [View at Publisher] [DOI] [Google Scholar]
17. Iorio R, Celenza G, Petricca S. Mitophagy: molecular mechanisms, new concepts on Parkin activation and the emerging role of AMPK/ULK1 Axis. Cells. 2021;11(1):30. [View at Publisher] [DOI] [PMID] [Google Scholar]
18. Eslimi Esfehani D, Oryan S, Khosravi M, Valizadegan F. Effect of fennel extract on the improvement of memory disorders in beta amyloid alzheimer model of male wistar rats. J Ilam Univ Med Sci. 2019;27(1):1-12. [View at Publisher] [DOI] [Google Scholar]
19. Bedford TG, Tipton CM, Wilson NC, Oppliger RA, Gisolfi CV. Maximum oxygen consumption of rats and its changes with various experimental procedures. J Appl Physiol Respir Environ Exerc Physiol. 1979;47(6):1278-83. [View at Publisher] [DOI] [PMID] [Google Scholar]
20. Wu C, Yang L, Li Y, Dong Y, Yang B, Tucker LD, et al. Effects of exercise training on anxious-depressive-like behavior in Alzheimer rat. Med Sci Sports Exerc. 2020;52(7):1456-69. [View at Publisher] [DOI] [PMID] [Google Scholar]
21. Monserrat Hernández‐Hernández E, Serrano‐García C, Antonio Vázquez‐Roque R, Díaz A, Monroy E, Rodríguez‐Moreno A, et al. Chronic administration of resveratrol prevents morphological changes in prefrontal cortex and hippocampus of aged rats. Synapse. 2016;70(5):206-17. [View at Publisher] [DOI] [PMID] [Google Scholar]
22. Fang EF, Hou Y, Palikaras K, Adriaanse BA, Kerr JS, Yang B, et al. Mitophagy inhibits amyloid-β and tau pathology and reverses cognitive deficits in models of Alzheimer's disease. Nat Neurosci. 2019;22(3):401-12. [View at Publisher] [DOI] [PMID] [Google Scholar]
23. Castellazzi M, Patergnani S, Donadio M, Giorgi C, Bonora M, Bosi C, et al. Autophagy and mitophagy biomarkers are reduced in sera of patients with Alzheimer's disease and mild cognitive impairment. Sci Rep. 2019;9(1):20009. [View at Publisher] [DOI] [PMID] [Google Scholar]
24. Cicero AF, Ruscica M, Banach M. Resveratrol and cognitive decline: a clinician perspective. Arch Med Sci . 2019;15(4):936-43. [View at Publisher] [DOI] [PMID] [Google Scholar]
25. Pagar KR, Khandbahale SV, Phadtare DG. The therapeutic potential of resveratrol: a review of clinical trials. Asian J Pharm Res. 2019;9(3):193-9. [View at Publisher] [DOI] [Google Scholar]
26. Weidling IW, Swerdlow RH. Mitochondria in Alzheimer's disease and their potential role in Alzheimer's proteostasis. Exp Neurol. 2020;330:113321. [View at Publisher] [DOI] [PMID] [Google Scholar]
27. Han Y, Wang N, Kang J, Fang Y. β-Asarone improves learning and memory in Aβ 1-42-induced Alzheimer's disease rats by regulating PINK1-Parkin-mediated mitophagy. Metab Brain Dis. 2020;35:1109-17. [View at Publisher] [DOI] [PMID] [Google Scholar]
28. Sarraf SA, Sideris DP, Giagtzoglou N, Ni L, Kankel MW, Sen A, et al. PINK1/Parkin influences cell cycle by sequestering TBK1 at damaged mitochondria, inhibiting mitosis. Cell Rep. 2019;29(1):225-35.e5. [View at Publisher] [DOI] [PMID] [Google Scholar]
29. Chen L, Qin Y, Liu B, Gao M, Li A, Li X, et al. PGC-1 α-mediated mitochondrial quality control: molecular mechanisms and implications for heart failure. Front Cell Dev Biol. 2022:10:871357. [View at Publisher] [DOI] [PMID] [Google Scholar]
30. Zhao N, Yan Q-W, Xia J, Zhang X-L, Li B-X, Yin L-Y, et al. Treadmill exercise attenuates Aβ-induced mitochondrial dysfunction and enhances mitophagy activity in APP/PS1 transgenic mice. Neurochem Res. 2020;45(5):1202-14. [View at Publisher] [DOI] [PMID] [Google Scholar]
31. Rostamzadeh F, Najafipour H, Aminizadeh S, Jafari E. Therapeutic effects of the combination of moderate-intensity endurance training and MitoQ supplementation in rats with isoproterenol-induced myocardial injury: The role of mitochondrial fusion, fission, and mitophagy. Biomed Pharmacother. 2024;170:116020. [View at Publisher] [DOI] [PMID] [Google Scholar]
32. Pantiya P, Thonusin C, Chunchai T, Pintana H, Ongnok B, Nawara W, et al. Long-term lifestyle intervention is superior to transient modification for neuroprotection in D-galactose-induced aging rats. Life Sci. 2023;334:122248. [View at Publisher] [DOI] [PMID] [Google Scholar]
33. Khodagholi F, Zareh Shahamati S, Maleki Chamgordani M, Mousavi MA, Moslemi M, Salehpour M, et al. Interval aerobic training improves bioenergetics state and mitochondrial dynamics of different brain regions in restraint stressed rats. Mol Biol Rep. 2021;48(3):2071-82. [View at Publisher] [DOI] [PMID] [Google Scholar]
34. Zhao N, Zhang X, Li B, Wang J, Zhang C, Xu B. Treadmill exercise improves PINK1/Parkin-mediated mitophagy activity against Alzheimer's disease pathologies by upregulated SIRT1-FOXO1/3 axis in APP/PS1 mice. Mol Neurobiol. 2023;60(1):277-91. [View at Publisher] [DOI] [PMID] [Google Scholar]
35. Das S, Mitrovsky G, Vasanthi HR, Das DK. Antiaging properties of a grape-derived antioxidant are regulated by mitochondrial balance of fusion and fission leading to mitophagy triggered by a signaling network of Sirt1-Sirt3-Foxo3-PINK1-PARKIN. Oxid Med Cell Longev. 2014:2014:345105. [View at Publisher] [DOI] [PMID] [Google Scholar]
36. Li H, Qin S, Liang Q, Xi Y, Bo W, Cai M, et al. Exercise training enhances myocardial mitophagy and improves cardiac function via Irisin/FNDC5-PINK1/Parkin pathway in MI mice. Biomedicines. 2021;9(6):701. [View at Publisher] [DOI] [PMID] [Google Scholar]
37. Zhou J, Wang Z, He Y, Luo X, Zhang W, Yu L, et al. Qiliqiangxin reduced cardiomyocytes apotosis and improved heart function in infarcted heart through Pink1/Parkin-mediated mitochondrial autophagy. BMC Complement Med Ther. 2020;20(1):203. [View at Publisher] [DOI] [PMID] [Google Scholar]
38. Lee J, Hong S-W, Kwon H, Park SE, Rhee E-J, Park C-Y, et al. Resveratrol, an activator of SIRT1, improves ER stress by increasing clusterin expression in HepG2 cells. Cell Stress Chaperones. 2019;24(4):825-33. [View at Publisher] [DOI] [PMID] [Google Scholar]
39. Wang B, Ge S, Xiong W, Xue Z. Effects of resveratrol pretreatment on endoplasmic reticulum stress and cognitive function after surgery in aged mice. BMC Anesthesiol. 2018;18(1):141. [View at Publisher] [DOI] [PMID] [Google Scholar]
40. Li Y, Xu S, Giles A, Nakamura K, Lee JW, Hou X, et al. Hepatic overexpression of SIRT1 in mice attenuates endoplasmic reticulum stress and insulin resistance in the liver. FASEB J. 2011;25(5):1664. [View at Publisher] [DOI] [PMID] [Google Scholar]
41. Um J-H, Park S-J, Kang H, Yang S, Foretz M, McBurney MW, et al. AMP-activated protein kinase-deficient mice are resistant to the metabolic effects of resveratrol. Diabetes. 2010;59(3):554-63. [View at Publisher] [DOI] [PMID] [Google Scholar]
42. Corpas R, Griñán-Ferré C, Rodríguez-Farré E, Pallàs M, Sanfeliu C. Resveratrol induces brain resilience against Alzheimer neurodegeneration through proteostasis enhancement. Mol Neurobiol. 2019;56(2):1502-16. [View at Publisher] [DOI] [PMID] [Google Scholar]
43. Solberg NO, Chamberlin R, Vigil JR, Deck LM, Heidrich JE, Brown DC, et al. Optical and SPION-Enhanced MR imaging shows that trans-stilbene inhibitors of NF-κB concomitantly lower alzheimer's disease plaque formation and microglial activation in AβPP/PS-1 transgenic mouse brain. J Alzheimers Dis. 2014;40(1):191-212. [View at Publisher] [DOI] [PMID] [Google Scholar]
44. Liao Z-Y, Chen J-L, Xiao M-H, Sun Y, Zhao Y-X, Pu D, et al. The effect of exercise, resveratrol or their combination on Sarcopenia in aged rats via regulation of AMPK/Sirt1 pathway. Exp Gerontol. 2017;98:177-83. [View at Publisher] [DOI] [PMID] [Google Scholar]
45. Broderick TL, Rasool S, Li R, Zhang Y, Anderson M, Al-Nakkash L, et al. Neuroprotective effects of chronic resveratrol treatment and exercise training in the 3xTg-AD mouse model of Alzheimer's disease. Int J Mol Sci. 2020;21(19):7337. [View at Publisher] [DOI] [PMID] [Google Scholar]
46. Benito-Cuesta I, Ordóñez-Gutiérrez L, Wandosell FJA. AMPK activation does not enhance autophagy in neurons in contrast to MTORC1 inhibition: different impact on β-amyloid clearance. Autophagy. 2021;17(3):656-71. [View at Publisher] [DOI] [PMID] [Google Scholar]
Send email to the article author
| Rights and permissions | |
 |
This work is licensed under a Creative Commons Attribution-NonCommercial 4.0 International License. |
